Your Cart is Empty

FEATURES / KEY INGREDIENTS:
-100% Pure Acetyl L-Carnitine
BENEFITS:
-Stimulant Free Fat Loss
-Weight Loss
-Cognitive Function
-Focus
GEN-TEC NUTRACEUTICALS source, blend and package 100% pure pharmaceutical grade ACETYL L-CARNITINE powder. ACETYL L-CARNITINE supplementation is ideal for individuals with a goal of improving mental focus while reducing subcutaneous bodyfat levels.
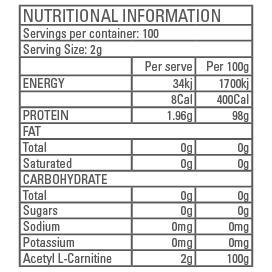
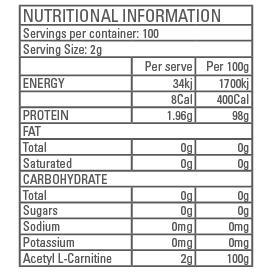
SUGGESTED USE: Mix 2g (1/2 tsp) of ACETYL L-CARNITINE in 150mL water and consume 20 minutes before aerobic or anaerobic exercise. Store in a cool, dry, dark place.
INGREDIENTS: 100% Pharmaceutical grade ACETYL L-CARNITINE, Silicon Dioxide (2%).
May contain traces of milk, soybeans, cereals containing gluten, tree nuts, sesame seeds and their products.
DISCLAIMER: Formulated Supplementary Sports Food. This product is not a sole source of nutrition and should be consumed in conjunction with a nutritious diet and an appropriate physical training or exercise program. Not suitable for children under 15 years of age or pregnant women. Should only be used under medical or dietetic supervision.
Made in Australia from local and imported ingredients.
Most customers receive their bodybuilding supplements in 3-5 business days!
Shipping to VIC - 3-5 business days
Shipping to NSW - 1-3 business days
Shipping to QLD - 3-5 business days
Shipping to NT/SA/TAS - 4-5 business days
Shipping to WA - 5-7 business days
Please note parcels going to PO Boxes, Parcel Lockers, Locked Bags or Australian Defence Addresses (via Australia Post) and bulky parcels going to WA/NT may take longer than normal deliveries.
DO YOU SHIP INTERNATIONALLY?
No, at this point in time we are unable to ship outside of Australia.
I HAVE ANOTHER QUESTION ABOUT SHIPPING!
If you haven’t found an answer to your question, please call our Customer Service team on 1300 792 609. We’ll be able to take your call from Monday – Friday 9.00am-5.00pm AEST.